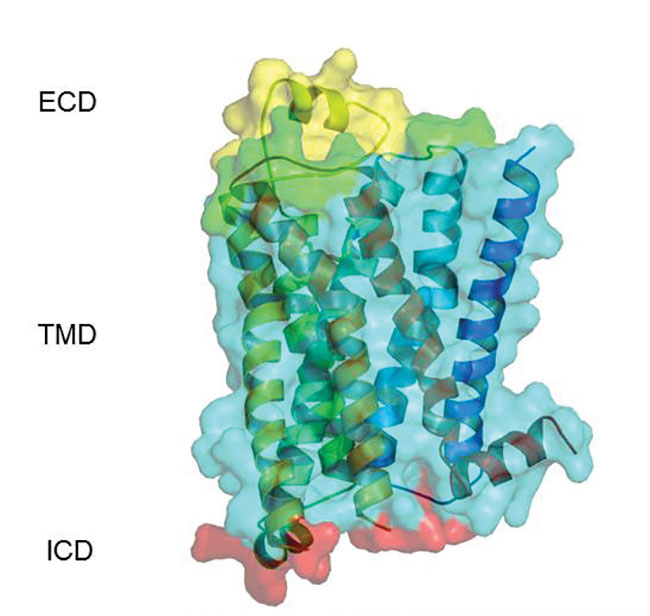
β1AR Crystal structure
depicting ECL2 in yellow
In part 1, we summarized the advances made in new approaches developed to address the challenges of antigen generation for targeting G protein–coupled receptors (GPCRs). We reviewed the antibody and biologics pipeline with progress highlighted by some interesting case studies on new targets (1). Here, we conclude by reviewing progress attained with other biologics.
Peptides Targeting G Protein–Coupled Receptors
More than 50 peptide-based therapeutic products are commercially available, but very few of them have been derived from recombinant display technology. In fact, only three peptide therapeutics targeting GPCRs have been discovered in that manner — one of which is ecallantide. Most others are either the endogenous agonist or a closely related analogue. However, in silico platforms also have been used for computational screenings. They are based on algorithms, such as Compugen’s peptide ligand discovery platform (2), which has identified potential therapeutic peptides, such as a LGR7 agonist and a FPRL1 agonist.
The current clinical pipeline consists of 128 peptide therapeutics, with about 50 of those targeting GPCRs. Global sales generated by approved peptide therapeutics targeting GPCRs exceeded US$5billion in 2011. GLP-1 and GLP-2 receptors are the most frequently targeted GPCRs, but overall, peptides target a wide range of GPCRs, including CXCR4, opioid, ghrelin, GnRH, oxytocin, LHRH, melanocortin, and PTH. A recent review thoroughly analyzed and summarized the peptide clinical pipeline and anticipated future directions for development of this drug class, such as peptide-drug conjugates that have taken key learnings from this area in antibody engineering (3). Table 1 lists examples of the main GPCR peptide targets.
The group of peptide modulators for GLP-1R (a family B GPCR) probably represent the largest group of therapeutic peptides targeting a GPCR target. Most of these aim for a drug regime of once-weekly dosing to facilitate patient compliance and convenience. Only three injectable peptides have been globally approved and marketed to date (exenatide or Byetta from AstraZeneca; exenatide-extended release formulation or Bydureon from AstraZeneca; and liraglutide or Victoza from Novo Nordisk). An oral weekly version of exenatide is also under development (Oramed Pharmaceuticals) with a pre-IND package submitted to the FDA in September 2013.
GLP-1R agonists continue to be of great interest with ever greater focus on further peptides gaining approval. Exenatide (and other peptides in development) is a synthetic derivative of exendin-4, which is isolated from the saliva of the Gila monster lizard, whereas others are stabilized analogues of GLP-1. By February 2013, one peptide agonist already had obtained approval in the European Union for T2D (lixisenatide). Further approvals are under way for T2D and erythropoietic protoporphyria (albiglutide and afamelanotide, respectively). Even dual agonists, such as RO6811135 (GLP-1R and GIPR), have been advanced to early clinical studies (clinical trial NCT01676584).
A number of alternative peptide structures have been investigated for their therapeutic effect. They use both agonist and antagonist modes of action, depending on the desired outcome. Examples include a truncated form of the RANTES peptide ligand that internalizes CCR5 and constrained or circular peptides — e.g., cyclohexapeptides such as octreotide, a synthetic peptide that is a somatostatin receptor agonist, developed for agromegaly and the oncology therapeutic area. Another example is pasireotide (which more closely mimics the activation of the receptor by natural ligands); it was developed for the treatment of Cushing’s disease and has a 40-fold increased affinity for somatostatin receptor 5 compared with other somatostatin analogues.
The development of small-molecule PAR2 ligands initially focused on modification of natural peptide agonists (4). Small molecule agonists have been identified through chemical library screens, and full PAR2 agonists have been described as having good stability but poor solubility. The first described nonpeptidic antagonist ENMD-1068 (selective for PAR2 over other PARs in vitro) had only very low potency. A more potent antagonist was reported in 2009, but it did not antagonize PAR2 activation by proteases such as trypsin. Anaturally derived ligand was reported to be toxic after oral administration in rats (5). Several other PAR2 antagonists have been described and were suggested to be potentially antiinflammatory, but little or no in vivo data were supplied in most cases. Studies on new antagonists have shown them to have antiinflammatory properties in vivo (6).
Yet another modality to small molecule ligands and peptide variants are pepducins (lipidated synthetic peptides) and worth mentioning here as alternative biologics. In early discovery stage, these have been designed to penetrate the cell membrane to target the intracellular domain of GPCRs to allosterically modulate receptor signaling (7). Advantages of using pepducins as potential therapeutics lie in their high selectivity for their targets and ease of modifications at specific amino acid residues. A small number of pepducins have been developed as allosteric modulators for PAR2 (8) but also FPR2 (9), S1P3 receptor (10), and CXCR4 (11). A PAR1 pepducin antagonist, PZ-128, is in development for treatment of coronary heart disease (clinical trial NCT01806077).
Bicycle Therapeutics is developing bi-macrocyclic constrained peptides, and Aileron Therapeutics is developing a constrained peptide format (Stapled Peptides) that is a proprietary stabilization technology that involves “stapling” peptides into an alpha-helical shape by means of hydrocarbon cross-linking chemistry. That format mimics the structure found at the interface of many protein–protein interactions. The first phase 1 clinical trial was initiated in the first half of 2013 for ALRN-5281 (a long-acting GHRH agonist) for treating orphan endocrine disorders, as well as broader patient populations involving a wide variety of metabolic/endocrine diseases.
Inventive methods for application as a therapeutic treatment other than directly dosing the patient targeting the GPCR have been devised. One example is a peptide corresponding to ECL2 of β1AR, which has been used for apheresis removal of β1AR agonist autoantibodies that cause the pathology seen in cardiomyopathyic diseases (12). A cyclic peptide, COR1, has now been designed to block anti-β1AR autoantibodies by mimicking the tertiary structure of ECL2 and demonstrated efficacy in a rat model of autoimmune cardiomyopathy (13). Subsequently, a phase 1 clinical study has confirmed safety in humans and good pharmacokinetic properties (14; clinical trial NCT 01043146).
Peptide modulators of GPCRs also are in development for nonpeptide receptors. One such example is the calcium-sensing receptor, where KAI-4169 is under development as a novel calcimimetic for the treatment of secondary hyperparathyroidism, originally by Kai Pharmaceuticals (15) (acquired by Amgen in 2012).
Technologies evaluated in antibody engineering also have been used to extend peptide half-life, including pegylation, albumination (albiglutide), linking to an AlbudAb (GLP-1/PYY dual combination, WO 2011/039096), and linkage of the peptide drug to an inert IgG scaffold known as a CoVX-body — such as CoVX096 that targets GLP-1 (16) — or to an Fc fragment known as a PeptiBody (17) (which extends the half-life by FcRn-mediated recycling). Currently no known peptibodies are targeting GPCRs in the public domain, but such peptides would offer a useful feature for half-life extension as does XTEN technology (Amunix Inc), which has recently been exemplified for GLP-2 (18).
Another innovative area showing great promise is the development of peptide–drug conjugates, either for drug delivery or radionuclide therapy. Such technology draws upon key learnings made from antibody engineering in which drug, linker, and antibody-targeting agent component technologies have been optimized.An established example of peptide receptor radionuclide therapy (PRRT) is the use of radiolabeled somatostatin analogues for tumor-cell killing. Similarly, cytotoxic compounds such as doxorubicin can be linked to hormone peptide analogues (e.g., LHRH, bombesin, and somatostatin) for targeting tumors that specifically express the corresponding receptors for those peptides, thereby providing the specificity of an antibody (19).
Other Scaffolds and Biologics
In addition to antibodies and peptides, a number of other scaffolds (e.g., nanobodies and other sources of biologics) are being used to target GPCRs. Nanobodies are naturally occurring heavy-chain only antibodies found in camelids that possess an exceptionally long CDR3 loop, where the nanobody is the reduced VH single domain (VHH). Particular interest is given to that loop because it is thought to confer an advantage in targeting deep clefts and grooves. A bitopic nanobody had been generated to CXCR4 for stem-cell mobilization but was not progressed beyond phase 1 clinical trials (20).
Shark IgNARs have a similar structure to a camelid heavy-chain only IgG (a long VH CDR3 loop) and can be engineered into smaller proteins such as shark VNARs, which have a very similar structure to nanobodies. Furthermore, they can be engineered to create an i-Body (AdAlta), which uses human protein as its framework and incorporates features of the shark antibody within that human scaffold. The technology has been applied to the identification of functional i-Bodies to CXCR4 (AdAlta, AU2012904915), with two antagonists and one agonist characterized so far.
Bovine antibodies also can contain long heavy-chain CDR3 loops that includes a trisulphide-bonded domain (21). They can be used as a therapeutic in much the same way as a VNAR, i-Body, or nanobody. Fabrus has reported the generation of a functional modulator to an undisclosed family B member.
Pieris initiated a collaboration with Pepscan to discover novel anticalins to GPCRs using Pepscan’s CLIPS technology, but no progress has been reported in public domain. Among the scaffold biologics, designed ankyrin repeat proteins (DARPins) have been isolated to the neurotensin receptor (NTS-1) by using ribosome display for cocrystallization (22).
Another emerging focus in the biologics area is the development of antisense technology. ISIS-GCGRRx is an antisense drug that targets the glucagon receptor (GCGR) to reduce the effects of glucagon for the treatment of T2D (which acts against the action of insulin and stimulates the liver to produce glucose) and improve glucose control. In addition, reducing GCGR produces more active glucagon-like peptide, or GLP-1, which assists in maintaining pancreatic function and enhancing insulin secretion. To date, a phase 1 study has been concluded with no clinically significant increases in lipids or blood pressure or hypoglycemic events. In addition, researchers reported an increase in total GLP-1, in agreement with previous preclinical observations. That antisense drug has now progressed to a phase 2 study in T2D patients with uncontrolled glucose levels despite being prescribed metformin (clinical trial NCT01885260).
Aptamers are globular RNA or DNA molecules. They can be developed to specifically bind a target and are identified through a process known as systematic evolution of ligands by exponential enrichment (SELEX). They offer potential as research tools with which to assess the role of cell surface receptors in normal tissue and disease pathology (23), as well as for cocrystallization to determine receptor structure by X-ray crystallography. GPCR-specific RNA aptamers have been generated to the neurotensin receptor (NTS-1) to explore their application for that purpose (24).
Challenges Remain
GPCRs are universally acknowledged as important therapeutic targets, but only recently has their potential as antibody targets come under greater scrutiny. We have determined that about 100 GPCR targets are suitable across a range of diseases (e.g., cancer, fibrosis, inflammation, respiratory, pain, metabolic) and commercially compelling as targets for antibodies and other biologics. Many technical challenges are encountered in the generation of antibodies to membrane proteins such as GPCRs and ion channels. However, developments in stabilizing receptors using mutagenesis or additives — as well as methods to overexpress receptors in cell systems — have enabled the identification of functional antibodies. For alternative scaffolds, future clinical candidates must ensure clear advantages over monoclonal antibody approaches based on unique scaffold attributes (e.g., pulmonary delivery and ocular applications). Wider acceptance will come with clinical success.
References
1 Koglin M, Hutchings CJ. Targeting G Protein–Coupled Receptors with Biologics for Therapeutic Use, Part 1. BioProcess Int. 12(6) 2014: 38–45.
2 Shemesh R, et al. Discovery and Validation of Novel Peptide Agonists for G Protein–Coupled Receptors. J. Biol. Chem. 283(50) 2008: 34643–34649.
3 Kasper AA, Reichert JM. Future Directions for Peptide Therapeutics Development. Drug Discov. Today 18(17–18), 2013: 807–817.
4 Yau MK, Liu L, Fairlie DP. Towards Drugs for Protease-Activated Receptor 2 (PAR2). J. Med. Chem. 56(19), 2013: 7477–7479.
5 Shao F, et al. Pharmacokinetic Study of Triptolide: A Constituent of Immunosuppressive Chinese Herb Medicine in Rats. Bio. Pharm Bull. 30(4), 2007: 702–707.
6 Adams MN. In Vitro and In Vivo Studies on Protease-Activated Receptor 2. PhD thesis. 2012. Queensland University of Technology, Brisbane, Queensland, Australia.
7 Carlson KE, McMurry TJ, Hunt SW III. Pepducins: Lipopeptide Allosteric Modulators of GPCR Signaling. Drug Discov. Today 9, 2012: 33–39.
8 Covic L, et al. Activation and Inhibition of G Protein–Coupled Receptors By Cell-Penetrating Membrane-Tethered Peptides. Proc. Natl. Acad. Sci. 99(2) 2002: 643–648.
9 Forsman H, et al. The Leukocyte Chemotactic Receptor FPR2, But Not the Closely Related FPR1, Is Sensitive to Cell-Penetrating Pepducins with Amino Acid Sequences Descending from the Third Intracellular Receptor Loop. Biochim. Biophys. Acta. 1833, 2013: 1914–1923.
10 Severino B, et al. Identification of a Pepducin Acting As a S1P3 Receptor Antagonist. J. Pept. Sci. 19(11) 2013: 717–724.
11 Quoyer J, et al. Pepducin Targeting the C-X-C Chemokine Type Receptor 4 Acts As a Biased Agonist Favouring Activation of the Inhibitory G Protein. Proc. Natl. Acad. Sci. 110(52) 2013: E5088–E5097.
12 Dandel M, et al. Immunoadsorption Therapy for Dilated Cardiomyopathy and Pulmonary Arterial Hypertension. Atherosclerosis Suppl. 14(1) 2013: 203–211.
13 Jahns R, et al. Targeting Receptor Antibodies in Immune Cardiomyopathy. Semin. Thromb. Hemost. 36(2) 2010: 212–217.
14 Münch G, et al. Administration of the Cyclic Peptide COR-1 in Humans (Phase 1 Study): Ex Vivo Measurements of Anti-β1-Adrenergic Receptor Antibody Neutralization and of Immune Parameters. Eur. J. Heart Failure 14(11) 2012: 1320–1329.
15 Walter S, et al. Pharmacology of AMG 416 (Velcalcetide), A Novel Peptide Agonist of the Calcium-Sensing Receptor for the Treatment of Secondary Hyperparathyroidism in Hemodialysis Patients. J. Pharmacol. Exp.Ther. 346(2) 2013: 229–240.
16 Woodnutt G, Violand B, North M. Advances in Protein Therapeutics. Curr. Opin. Drug Discov. Dev. 11(6) 2008: 754–761.
17 Shimamoto G, et al. Peptibodies: A Flexible Alternative Format to Antibodies. MAbs 4(5) 2012: 586–591.
18 Podust VN, et al. Extension of In Vivo Half-Life of Biologically Active Peptides via Chemical Conjugation to XTEN Protein Polymer. Protein Eng. Des. Sel. 26(11) 2013: 743–753.
19 Thundimadhathil J, et al. Cancer Treatment Using Peptides: Current Therapies and Future Prospects. J. Amino Acids 2012: 967347.
20 Jähnichen S, et al. CXCR4 Nanobodies (VHH-Based Single Variable Domains) Potently Inhibit Chemotaxis and HIV-1 Replication and Mobilize Stem Cells. Proc. Natl. Acad. Sci. 107(47) 2010: 20565–20570.
21 Wang F, et al. Reshaping Antibody Diversity. Cell 153(6) 2013: 1379–1393.
22 Milovnik P, et al. Selection and Characterization of DARPins Specific for the Neurotensin Receptor 1. Protein Eng. Des. Sel. 22(6) 2009: 357–366.
23 Cibiel A, Dupont DM, Ducongé F. Methods to Identify Aptamers Against Cell Surface Biomarkers. Pharmaceuticals 4 2011: 1216–1235.
24 Daniels D, et al. Generation of RNA Aptamers to the G-Protein Coupled Receptor for Neurotensin, NTS-1. Anal. Biochem. 305(2) 2002: 214–226.
Markus Koglin, PhD, is associate director, protein engineering; and corresponding author Catherine J. Hutchings is principal scientist, antibody alliance management and strategic partnering, at Heptares Therapeutics Ltd, BioPark, Broadwater Road, Welwyn Garden City, Herts AL7 3AX; 44-1707358698; fax 44-1707358640; cath.hutchings@heptares.com.
The HEPTARES name, the logo, and STAR are trademarks of Heptares Therapeutics Ltd.